记忆T细胞可以介导快速有效的免疫反应,提供长期的免疫保护。根据其在体内的特殊功能和分布,记忆T细胞可以简单地分为两大类:循环记忆T细胞和组织驻留记忆T细胞(TRM)。循环记忆T细胞包括中枢记忆T细胞(TCM)和效应记忆T细胞(TEM),它们通过淋巴和血液调节机体。而TRM细胞仅限于外周组织,最近被认为是外周器官中重要的保护性介质。
结合转录和表型分析发现,TRM细胞的典型特征为高表达组织驻留相关分子,如CD69,以及下调组织输出的相关基因,包括鞘氨醇I磷酸受体1(S1PR1)和C-C趋化因子受体7(CCR7)。受到刺激后,TRM细胞迅速释放效应分子,启动免疫防御,并诱导募集其他免疫细胞,如调节性T细胞(Treg)、中性粒细胞、自然杀伤细胞和树突状细胞。TRM细胞广泛存在于肝脏、肺、肠和皮肤等器官中。根据细胞表面共受体,TRM细胞在不同的组织部位分为CD8+和CD4+亚群。在这些器官中,肝脏是维持体内平衡的关键,也是许多代谢和免疫过程的核心,常面临病原体侵袭和食物源性抗原的双重挑战。在暴露于抗原时,肝脏TRM细胞参与到适应性细胞防御,与其他细胞共同发挥作用消除病原体。本文总结了TRM细胞在慢性肝病中的表型和它们在这些疾病中的潜在治疗靶点,以期有助于慢性肝病的诊断和治疗。
1TRM细胞
TRM细胞驻留在组织内,具有不同的代谢特征和转录机制(表1)。TRM细胞通常以其定位和效应功能为特征,而定位和效应功能由表面标志物及转录特征决定。研究发现,尽管TRM细胞具有组织特异性,但它们也存在统一的特征,能够表达一些黏附分子,如CD69、CD103 和CD49a,并关闭一些可以促进细胞离开组织的基因,如Krüppel样因子2(KLF-2)和S1P受体,以及能上调调控T细胞驻留的组织特异性分子。
凝集素CD69是调控细胞在组织驻留的关键,在大多数不同的TRM细胞亚群中组成性表达。当暴露于抗原或促炎介质时,由于KLF-12的下调,CD69的表达显著上调。同时,CD69作为S1P1的拮抗剂,通过介导S1P1的内化和降解促进肝脏TRM细胞的驻留,并下调S1PR1介导的组织输出。因此,CD69的主要作用可能是限制TRM细胞从肝脏排出到血管和淋巴结。
CD103是整合素αEβ7的α组分,在转化生长因子β(TGF-β)的作用下表达水平显著升高。钙黏蛋白E是一种连接上皮细胞的黏附蛋白。作为钙黏蛋白E的受体,TRM细胞上的CD103可与钙黏蛋白E结合,这对TRM细胞的定位、黏附和保留至关重要。此外,CD49a作为TRM细胞的另一个表面标志物,是整合素α1β1的α链,其与整合素β1结合形成的异二聚体VLA-1,可以促进整合素β1与Ⅳ型胶原结合,导致驻留细胞在上皮的维持。T细胞活化后,CD49a水平上调。据报道,CD49a的阻断和缺失会导致TRM细胞的减少。
在TRM细胞的形成和维持过程中,许多转录因子发挥了调控作用,包括B细胞诱导成熟蛋白1(BLIMP1)、T细胞中BLIMP1的同源物(HOBIT)、RUNT相关转录因子3(RUNX3)、E类基本螺旋环螺旋蛋白40(BHLHE40)和T盒转录因子21(TBX21/T-bet)。TRM细胞的转录谱是其拥有独特特征的基础。受到抗原刺激时,HOBIT与BLIMP1表达上调,在外周组织中介导TRM细胞的发育。同时,HOBIT和BLIMP1的共表达可抑制TRM细胞中CCR7、KLF-2及S1PR1的表达,而CCR7有助于细胞向外周淋巴组织迁移,S1PR1和KLF-12对淋巴细胞的组织抑制至关重要,因此,HOBIT与BLIMP1结合可抑制与组织排出相关的基因和标志物的表达,从而保留TRM细胞。RUNX3可以下调与循环T细胞发育有关的基因,上调包括CD103在内的驻留相关分子。BHLHE40可诱导应激反应蛋白的表达,以促进TRM细胞在应激状态下的效应功能。T-bet有助于提高IL-15受体和CD18的水平,使TRM细胞能够长期存活。所有这些因子都被证明对TRM细胞的分化和维持具有不可或缺的影响。
除了表面标志物和转录因子外,趋化因子和趋化因子受体已被报道对外周组织中TRM细胞的驻留及效应功能至关重要。尽管它们在不同组织的TRM细胞上的表达具有高度的特异性,但据报道,TRM细胞的维持及其效应功能的发挥,依赖于持续性趋化因子信号转导的调控。CXCR3与CXCR6广泛表达于不同组织来源的TRM细胞表面。研究证实,这些受体通过识别单核细胞、肝窦内皮细胞及成纤维细胞分泌的CXCL9、CXCL10、CXCL11与CXCL16等配体,协同调控TRM细胞的驻留与维持。
2肝脏中的TRM细胞
尽管不同组织中的TRM细胞具有某些共同特征,但其表型特征、组织驻留机制及效应功能仍表现出显著的异质性。肝脏中的TRM细胞通过独特的免疫调节机制,在病原体清除和自身抗原介导的免疫反应中发挥关键作用。
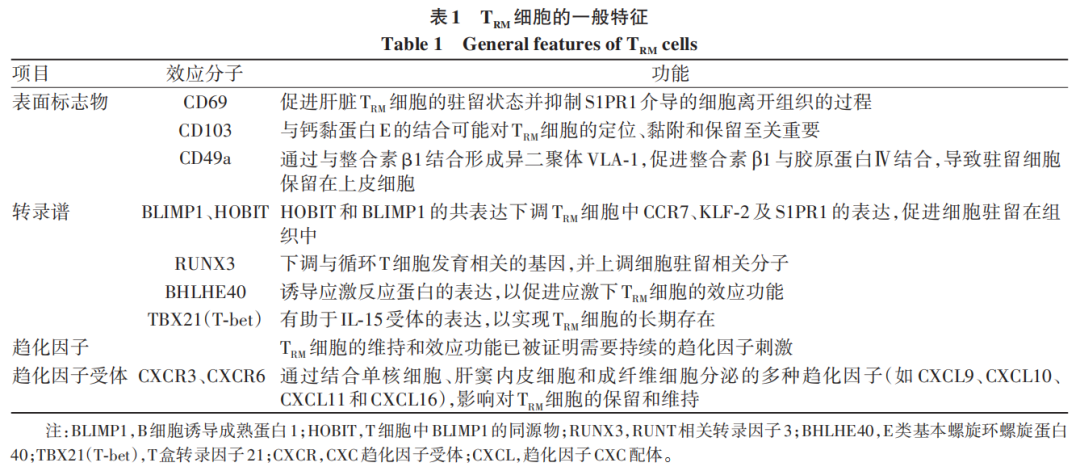
据报道,肝脏TRM细胞的生成和维持基础在于其表型特征,包括表面标志物和转录谱(图1)。与其他器官内的TRM细胞类似,肝脏TRM细胞上调黏附分子和趋化因子受体,包括CD69、CD103、CD49a、CXCR3和CXCR6,并下调组织排出的基因水平,如KLF-2和S1P1。凝集素CD69通过下调KLF-2和S1P1的水平,影响肝脏TRM细胞的保留。此外,在人类肝脏中,根据CD103的表达与否,存在两种功能不同的TRM细胞亚群,其中CD69+CD103+亚群作为特异于抗原的细胞毒性T细胞,较CD69+CD103-亚群显示出更强的效应功能。人与小鼠肝脏TRM细胞中CD103的表达也存在一些差异。其他肝脏特异性归巢标志物,包括CD11a和淋巴细胞功能相关抗原1(LFA-1),可能是导致小鼠肝脏TRM细胞驻留的原因。已有研究发现,LFA-1与细胞间黏附分子1结合后,可介导CD103细胞在微血管中移动。
注: IL-15R,白细胞介素15受体;IL-15,白细胞介素15; TGF-βR,转化生长因子β受体;TCR,T细胞抗原受体;MHCⅠ,主要组织相容性复合体Ⅰ类分子。
图1 肝脏CD8+ TRM细胞的表型和代谢特征
TRM细胞的代谢谱系具有明显的组织特异性。肝脏TRM细胞的分化与稳态维持呈现出独特的能量代谢特征,氧化磷酸化和线粒体功能被证实是其发育成熟及长期驻留的关键调控因素。一般来说,高度活跃的细胞主要通过糖酵解途径提供能量,而静止细胞则通过氧化磷酸化和脂肪酸氧化途径提供能量。哺乳动物雷帕霉素靶蛋白(mTOR)在调节T细胞增殖和活化方面具有重要作用,其诱导葡萄糖消耗的功能可以支持TRM细胞的生成。脂肪酸β-氧化已被证明是CD8+TRM细胞发育和成熟的关键调控因子。细胞内分子脂肪酸结合蛋白(FABP)对脂肪酸代谢至关重要,研究表明,肝脏TRM细胞上调FABP1而不表达FABP5,当FABP1缺失时,肝脏TRM细胞功能受损,而FABP1的重新表达可使其功能恢复。P2X嘌呤受体7(P2RX7)是一种胞外核苷酸受体,在细胞外三磷酸腺苷(eATP)的刺激下可促进线粒体止血并维持T细胞代谢;P2RX7还可通过钙调磷酸酶信号传导上调TGF-β受体,支持TRM细胞的产生和维持。
3肝脏TRM细胞在慢性肝病中的作用
3.1 病毒性肝炎
病毒性肝炎是由多种嗜肝病毒引起的传染性疾病,包括乙型肝炎病毒(HBV)、丙型肝炎病毒(HCV)、丁型肝炎病毒(HDV)等,可导致肝细胞损伤,甚至肝功能衰竭。肝脏TRM细胞参与肝脏慢性病毒感染的抗病毒免疫反应。
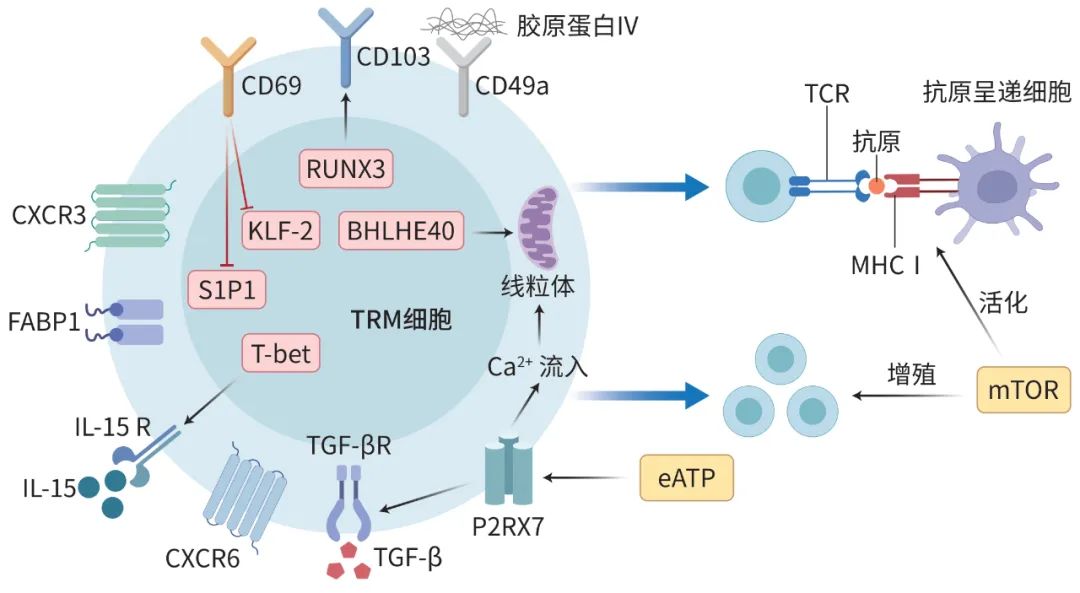
Pallett等首次发现,在HBV感染后的肝脏炎症过程中,经过T细胞抗原受体(TCR)和细胞因子介导的连续刺激,肝内记忆T细胞更有可能获得TRM表型。CD8+TRM细胞可在肝脏中存在数年之久,在康复的HBV患者中仍能检测到,这表明CD8+TRM细胞具有长期效应功能。此外,慢性HBV感染者的肝脏中存在大量CD69+CD4+TRM细胞,这与炎症的进展有关。在乙型肝炎进展过程中,肝内其他细胞的程序性死亡-配体1(PD-L1)表达上调,与TRM细胞上的程序性死亡受体1(PD1)结合诱导促炎症TRM细胞反应。同时TRM细胞强烈表达肿瘤坏死因子α(TNF-α)和IL-2,以消除感染的肝细胞。Wu等研究表明,干扰素γ(IFN-γ)I+Th1细胞通过驱动M1巨噬细胞的功能极化,激活肝内TRM细胞,促进乙型肝炎表面抗原(HBsAg)消失。此外,CD8+TRM细胞的数量与HBV病毒载量呈负相关,这表明肝脏TRM细胞具有调控感染细胞中病毒复制的功能。同时,表达CD44和LFA-1的Asialo GM1(AGM1)阳性CD8+TRM细胞已被证实对HBV的免疫清除至关重要。因此,增加肝脏TRM细胞数量可能是慢性HBV感染的潜在治疗方法(图2a)。
注: a,病毒性肝炎中的肝脏TRM细胞;b,MAFLD中的肝脏TRM细胞;c,自身免疫性肝病中的肝脏TRM细胞;d,肝硬化中的肝脏TRM细胞。PANX1,泛连接蛋白1;CCL5,CC类趋化因子配体5;GzmB,颗粒酶B。
图2 不同慢性肝病中的肝脏TRM细胞
慢性HCV感染时,人体内出现大量的CD4+和CD8+ TRM细胞;同样地,在HCV感染后恢复的动物模型中,CD69+CD8+T细胞大量增加。这些结果表明TRM细胞在控制HCV感染中起重要作用。
此外,HDV感染引起的炎症和疾病发展与CD8+TRM细胞的抗原非特异性激活密切相关。HDV通常会逃避CD8+T细胞的识别,从而导致不良的预后。CD8+TRM细胞会上调先天样活化受体自然杀伤细胞家族2成员D(NKG2D)的表达,从而诱导免疫介导的病毒控制(图2a)。总之,肝脏TRM细胞对清除慢性病毒感染和改善疾病预后至关重要。
3.2 代谢相关脂肪性肝病(MAFLD)
当机体有代谢综合征、高血压和2型糖尿病时,肝脏表现为MAFLD。在MAFLD中,CD8+TRM细胞已被证明在肝纤维化过程中发挥双重影响,它既是肝纤维化的驱动因素,也是消退因素。这些肝脏CD8+TRM细胞的特点是叉头框蛋白O1(FOXO1)转录因子活性低,CXCR6表达上调,这使得它们容易受到代谢活动的刺激并引发自身免疫性反应。此外,肝脏CD8+TRM细胞还可以通过产生促炎细胞因子,促进TRM细胞的生成(图2b)。一项研究表明,TRM通过免疫受体NKG2D激活和分泌IL-17A,使肝细胞产生趋化因子,将促炎细胞招募到肝脏,最终导致非酒精性脂肪性肝炎(NASH)和肝纤维化。然而也有报道称肝脏CD8+TRM细胞参与了NASH患者肝纤维化的缓解,研究表明,这些CD8+TRM细胞能够以CCR5依赖的方式吸引肝星状细胞,并引起FasL-Fas介导的活化肝星状细胞的凋亡。
3.3 自身免疫性肝病
自身免疫性肝病是一组以自身免疫异常为特征的疾病,包括自身免疫性肝炎(AIH)、原发性硬化性胆管炎(PSC)、原发性胆汁性胆管炎(PBC)和重叠综合征(OS)。
AIH是一种由过度活跃的T细胞介导的慢性炎症性肝病。多项研究揭示了CD8+TRM细胞在AIH进展中的重要作用。在AIH患者的肝脏中,CD69+CD103+CD8+TRM细胞增加,这与血清丙氨酸转氨酶、免疫球蛋白G水平、肝脏炎症和纤维化阶段呈正相关,而糖皮质激素能直接抑制TRM细胞的扩增。此外,AIH患者的肝脏TRM细胞表达更高水平的IL-15和TGF-β来诱导TRM细胞的产生。同时,AIH患者的肝细胞可通过上调CD103的配体钙黏蛋白E的水平来促进CD8+TRM细胞的驻留。钙黏蛋白E在胆管细胞中也被广泛表达,Zimmer等研究证实,CD8+TRM细胞比CD103-TRM细胞更接近胆管上皮细胞,这表明CD8+TRM细胞可能是胆管损伤发病机制的一部分。有报道称,PD-1高表达的CD8+TRM细胞可介导PBC中胆管上皮细胞的细胞毒性。Poch等研究显示PSC患者的胆管炎症中存在CD4+TRM细胞(图2c)。此外,最近Li等研究发现,衣康酸能影响DNA去甲基化以影响TRM相关基因表达,进而通过抑制CD103+TRM细胞,减轻PSC患者的肝脏和胆管损伤。
3.4 肝硬化
肝硬化是多种因素持续或反复作用下形成的肝脏弥漫性损伤,表现为广泛的肝细胞变性、坏死和纤维组织弥漫性增生。在致病因子作用下受损的肝细胞通过释放大量炎症因子,招募免疫细胞表达细胞因子,进而激活肝星状细胞,最终增加了细胞外基质的数量。肝星状细胞是肝纤维化的主要效应细胞,它们能转分化为产生胶原的肌成纤维细胞。多项研究表明,TRM细胞对肝硬化的发展有着不可忽视的影响。在肝硬化患者中,肝脏CD69+CD103-CD8+TRM亚群被活化,并表现出对缺氧诱导因子2α(HIF-2α)依赖性的细胞因子合成增强。此外,Koda等研究表明,CD8+TRM细胞可通过CCR5依赖性趋化吸引机制吸引造血干细胞,进而通过FasL/Fas依赖性凋亡直接对造血干细胞产生细胞毒性作用(图2d)。随着CD8+T细胞的减少,肝硬化和肝内炎症的消退停止,这表明CD8+T细胞通过增强造血干细胞的自噬作用来抑制肝硬化。
3.5 肝移植
肝移植是治疗终末期肝病的最后和唯一有效的手段。供体来源的TRM细胞已被证明在同种异体肝移植中持续存在,它们的丰度可能是不良事件发生率升高和器官存活率降低的原因。然而,还需要进一步研究肝脏供体来源的TRM细胞与不良事件发生率之间的联系。研究显示,2%~6%的CD8+T细胞在移植11年后出现供体来源的TRM表型,同时在肝脏和血液中可检测到TRM表型受体的T细胞数量较少。此外,供体来源的TRM细胞可通过下调某些TRM细胞的标志物,迁移到外周淋巴结。最近Li等研究发现CD8+TRM细胞与肝移植的急性反应及排斥水平密切相关,在同基因移植小鼠耐受模型中,CD8+TRM细胞在急性期显著上调,而在耐受期则下调,这揭示了TRM细胞在肝移植术后介导排斥反应的新机制。在未来,研究应更多地关注TRM细胞的特定靶点和信号传导位点,以改善移植的临床排斥反应。
4肝脏TRM细胞的潜在治疗靶点
4.1 核苷酸二磷酸X水解酶1(NUDT1)
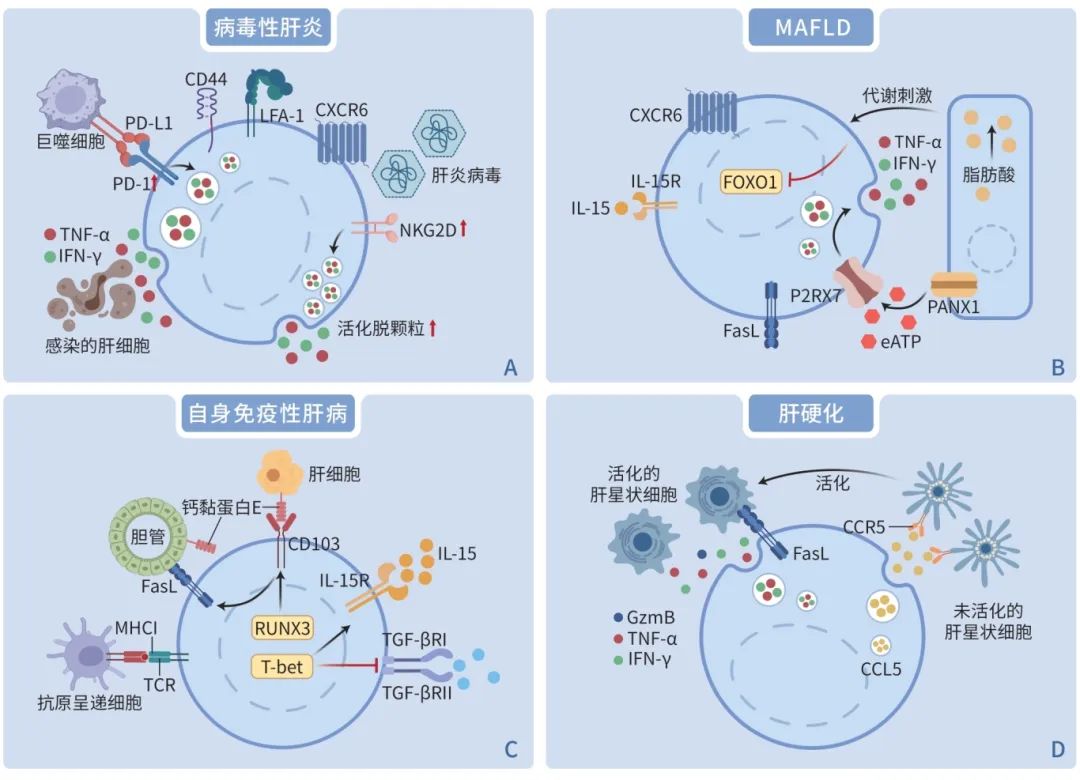
研究表明,持续的抗原刺激会限制T细胞线粒体的氧化能力。然而,线粒体活性的增加是各种组织中TRM细胞长久存在的必要条件,与此同时线粒体活性的增加可能导致产生过量的活性氧,进而造成DNA的氧化损伤。Huang等研究发现,肝脏CD103+TRM细胞会上调NUDT1的表达。作为一种核苷酸降解酶,NUDT1不仅可增强对氧化应激的耐受性,还可以通过抑制多聚ADP核糖聚合酶1(PARP1)的过度激活来增强TGF-β-Smad信号通路(图3)。Dudek等研究显示,高表达NUDT1的CD103+TRM细胞更靠近受损的小叶间胆管,同时表达更多的细胞因子。在PBC小鼠模型中,敲除或阻断NUDT1可减少CD103+TRM细胞,可以减轻门静脉的炎症。尽管尚未有明确证据表明NUDT1的活性与T细胞驱动的自身免疫中活性氧的生成有关,但NUDT1可能是调控TRM细胞的潜在靶点之一。
注: PRDM1,含PR域蛋白1;NR3C1,核受体亚家族3C组成员1;GR,糖皮质激素受体;TETs,10-11易位家族。
图3 肝脏TRM细胞作为免疫疗法的潜在靶点
4.2 P2RX7
eATP是真核生物用于检测细胞损伤的信号之一。炎症和损伤期间释放的eATP可通过嘌呤能受体P2RX7激活先天性免疫。研究发现,P2RX7可促进线粒体稳态和新陈代谢功能,有助于维持和生成CD8+TRM细胞。此外,CD8+TRM细胞会上调P2RX7的表达以增强对eATP的识别,进而导致肝细胞凋亡(图3)。P2XR7已被认为是可用于慢性疼痛治疗的药理学靶点,也是未来免疫疗法的潜在靶点。
4.3 糖皮质激素受体(GR)
GR由核受体亚家族3C组成员1(NR3C1) 基因编码,是核受体超家族的一员。它能与细胞质中的糖皮质激素结合,转移到细胞核中,并与DNA或其他蛋白质相互作用,发挥多种生物学功能。研究表明,糖皮质激素可以通过抑制CD8+TRM细胞的增殖来缓解AIH。NR3C1可与含PR域蛋白1(PRDM1)的启动子结合,而PRDM1可编码BLIMP1,BLIMP1在CD8+TRM细胞的分化和维持中起主要作用。GR可以下调PRDM1的表达,从而抑制CD8+TRM细胞(图3)。这些研究表明,未来可通过GR作用于CD8+TRM细胞,进而治疗慢性肝病。
4.4 CCR5
趋化因子及趋化因子受体已被证明对组织内TRM细胞的维持和效应功能至关重要。在肝硬化过程中,造血干细胞的CCR5表达逐渐减弱,然后在肝硬化缓解时逐渐恢复。研究证明,CD8+T细胞通过依赖CCR5的趋化作用对造血干细胞产生细胞毒性作用,从而缓解肝硬化(图3)。此外,CCR5还能吸引CD69HICD4+TRM细胞到达门静脉区发挥免疫效应。因此,靶向CCR5可能对肝硬化有治疗作用。
4.5 RUNX3
RUNX3是TRM细胞分化和维持稳态不可替代的调节因子,它不仅驱动CD8+T细胞在组织中驻留的程序,还与其他因子协同促进细胞毒性基因的表达。有报道称,衣康酸可通过干扰CD8+T细胞中的RUNX3基因启动子区的去甲基化,显著降低致病性CD103+TRM细胞的诱导和效应功能,缓解PBC的症状。此外,RUNX3可以影响细胞中CCR3和CCR5的表达,间接发挥其免疫作用(图3)。因此,在未来RUNX3有望成为改善肝损伤的治疗靶点。
4.6 磷酸化蛋白53(P53)
JAK/STAT信号通路对于T细胞的阴性选择和维持外周T细胞的正常表型及功能具有重要的调节作用。信号转导及转录激活蛋白3(STAT3)的过度激活与免疫抑制和转化有关,可通过一系列信号转导过程调控Th17细胞的分化。P53是一种转录因子,在调节细胞周期、细胞凋亡和基因组稳定性方面起着关键作用。在最近发表的一项研究中,Zhou等发现,P53作为JAK3/STAT3通路的下游分子,受免疫反应基因1(IRG1)/衣康酸的调控,诱导CD8+TRM细胞凋亡,从而缓解刀豆蛋白A诱导的AIH。因此,P53是治疗自身免疫性肝病的潜在关键靶点,有可能减轻自身免疫性肝病造成的肝损伤。
5小结与展望
TRM细胞在肝脏中的维持及其效应功能揭示了其在慢性肝病中的潜在作用。目前的研究已经在考虑利用TRM细胞对肿瘤进行免疫治疗,利用TRM细胞参与的信号通路治疗疾病,以及开发TRM细胞相关疫苗。然而,肝脏TRM细胞的免疫学和慢性肝病的免疫疗法仍有待研究。
多项研究表明,缺乏活性TGF-β时,抗原特异性TRM 细胞优于非特异性TRM细胞,两者间的竞争在肝病治疗中至关重要。而Treg细胞可通过提升TGF-β的生物利用度,影响TRM细胞生成。对肺纤维化的研究显示,牛黄脱氧胆酸可抑制TRM 样Treg细胞增殖,促进肺组织TEM细胞扩张。同样地,Rakhra等利用白蛋白转运疫苗以诱导黏膜TRM细胞,这依赖于交叉呈递的树突状细胞的激活。未来研究应结合其他免疫细胞与肝脏TRM细胞的相互作用,探索新药的开发。
随着对慢性肝病发病机制的深入研究,肝脏TRM细胞的潜力将被逐步挖掘。由于新陈代谢和转录机制的不同,肝脏中的TRM细胞与其他组织中的TRM细胞之间的区别以及在慢性肝病中的复杂作用逐渐显现。肝脏TRM细胞的相关通路可作为各种慢性肝病的免疫治疗靶点,让患者获得更准确、更持久的治疗。
全文下载
https://www.lcgdbzz.org/cn/article/doi/10.12449/JCp50526
